2022年10月31日,本实验室张智红教授团队,应用活体光学分子成像技术可视化研究胞内抗原诱导的免疫清除肝转移肿瘤的效应与机制,揭示了在肝脏转移形成过程中肿瘤细胞内ROS-caspase-3-GSDME通路活化是触发胞内抗原诱导的免疫清除肝转移瘤的关键步骤。研究成果以“Intravital molecular imaging reveals that ROS-caspase-3-GSDME-induced cell punching enhances humoral immunotherapy targeting intracellular tumor antigens”为题发表在Theranostics 12(17):7603-7623, 2022。
基于肿瘤抗原的肿瘤疫苗研制是肿瘤免疫治疗的重要方向之一。肿瘤抗原分为细胞胞内抗原和胞外抗原。由于细胞膜的屏障作用,胞内抗原无法直接被抗体识别与结合。靶向胞内抗原的肿瘤免疫治疗的关键之一是,改变细胞膜的通透性,提高抗体对胞内抗原的识别与结合。肿瘤细胞的分子信号变化与细胞膜的通透性有着密切关系。然而,目前对于活体微环境下肿瘤细胞膜通透性受哪些因素的影响及其分子调控机制仍不十分清楚。因此,通过活体光学分子成像观察肿瘤细胞内的分子信号变化,研究促进肿瘤细胞胞内抗原释放并与血液中抗体结合的分子机制,将有助于发展基于胞内抗原的肿瘤疫苗的开发与应用。
该研究以四聚体远红荧光蛋白(tfRFP)作为胞内抗原模型,构建了tfRFP皮下免疫小鼠后的tfRFP-B16肿瘤肝转移模型。结果显示,在未免疫组(Non-IM)中,小鼠肝脏内可检测到明显的tfRFP-B16荧光信号,而在tfRFP免疫组(tfRFP-IM)中,小鼠肝脏未能检测到tfRFP-B16荧光信号(图 1A-C)。同一批样本的肝脏实物图显示,在未免疫组肝脏体积出现不同程度的增大,呈现大小不一的转移灶点,而tfRFP组的肝脏没有明显异常(图 1D)。这些结果表明,tfRFP-B16细胞经脾注射能够形成明显的肿瘤肝转移灶,而经胞内抗原tfRFP皮下免疫的小鼠能够完全抑制或清除tfRFP-B16细胞的肝转移。
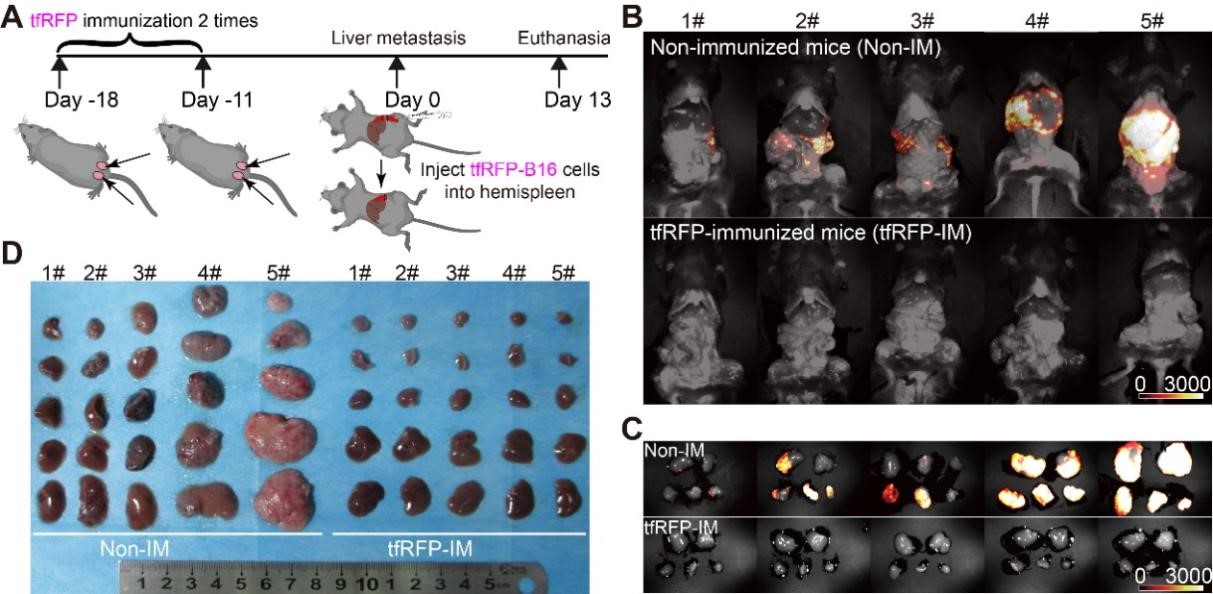
图 1. tfRFP尾根部皮下免疫抑制tfRFP-B16黑色素瘤细胞的肝转移。(A)tfRFP免疫和肝转移模型建立的流程图。(B)tfRFP-B16肿瘤细胞经脾注射后第13天小鼠整体荧光成像图。(C)tfRFP-B16肿瘤细胞经脾注射后第13天小鼠肝脏整体荧光成像图。(D)肿瘤经脾注射后第13天小鼠肝脏实物图。
为了探究tfRFP免疫小鼠体内的特异性免疫效应清除tfRFP-B16肝转移瘤的时空信息,研究者运用活体肝脏长时程光学显微成像窗口模型对tfRFP-B16肝转移过程进行连续数天观测。活体成像结果表明,tfRFP免疫之后,tfRFP-B16细胞经脾注射进入肝脏后会先定植与生长,然后才被逐渐清除掉,清除肿瘤细胞的关键时间点是在肿瘤细胞转移后的前7天。
研究者提取了tfRFP免疫小鼠的腹股沟引流淋巴结,进行免疫荧光染色分析。结果显示,皮下免疫的tfRFP抗原蛋白主要蓄积在腹股沟淋巴结的生发中心,并被滤泡树突状细胞摄取 (图 2A)。血清中可检测到大量的anti-tfRFP特异性抗体(图 2B)。肝脏活体光学成像观察到tfRFP免疫组的肿瘤转移灶内有tfRFP+微颗粒形成(图 2D-E)。对tfRFP免疫小鼠肝转移组织切片进行免疫荧光染色,成像结果显示,tfRFP免疫组肝脏肿瘤转移灶内的tfRFP+微颗粒是IgG阳性的,且没有细胞膜包裹(DiI阴性)(图 2F)。上述研究结果表明,tfRFP皮下免疫诱导所产生的抗体能够导致tfRFP-B16细胞产生tfRFP+微颗粒,该tfRFP+微颗粒是抗原抗体复合物。
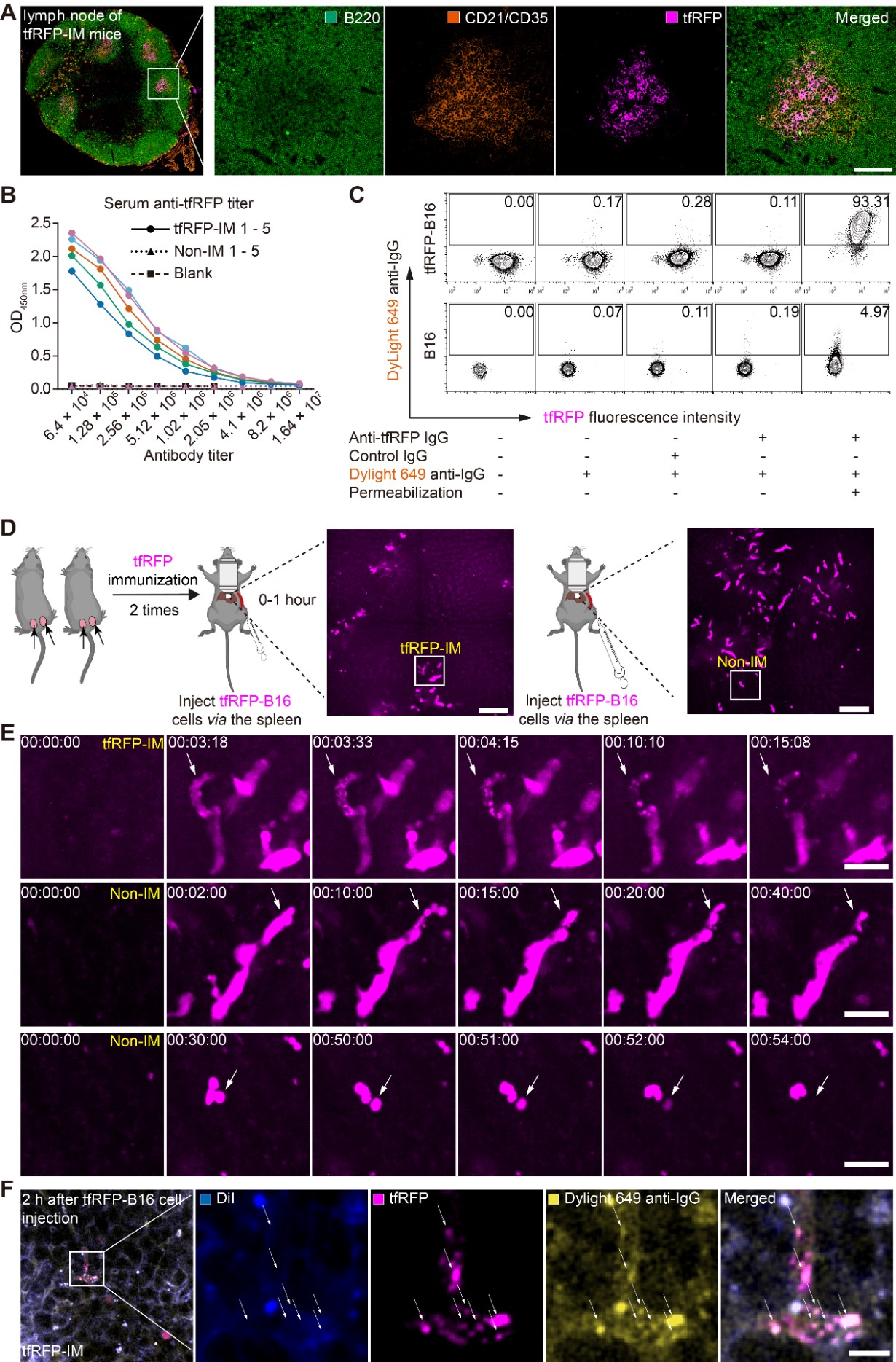
图 2. tfRFP 免疫诱导的体液免疫反应导致 tfRFP-B16进入肝脏的早期阶段形成tfRFP+抗原抗体复合物。(A)tfRFP免疫小鼠腹股沟淋巴结切片免疫荧光图。标尺:100 μm。(B)对比tfRFP免疫小鼠血清和未免疫小鼠血清anti-tfRFP IgG抗体滴度。(C)流式分析tfRFP-B16细胞在没有固定破膜或固定破膜条件下孵育anti-tfRFP IgG后细胞的IgG结合量。(D)经脾注射的tfRFP-B16细胞进入肝血窦过程的活体成像。标尺:100 μm。(E)对应A图中的小方框放大的时间序列图像,上排为tfRFP免疫小鼠中的tfRFP-B16细胞破裂形成tfRFP+微颗粒过程,下排为未免疫小鼠的tfRFP-B16细胞破裂消失过程。标尺:30 μm。(F)tfRFP免疫小鼠中的tfRFP+微颗粒为抗原抗体复合物。标尺:10 μm。
接着,研究者运用活体光学分子成像揭示在早期肿瘤细胞定植肝脏过程中的分子事件。将表征ROS和Caspase-3的MLS-HyPer7-B16细胞和tfRFP-C3-B16细胞混合后经脾注射到ROSAmT/mG小鼠中,观察肿瘤细胞到达肝脏时的ROS和Caspase-3的变化(图 3A)。结果显示,肿瘤细胞在经脾注射2 min内就能滞留于肝脏血窦中,MLS-HyPer7-B16 细胞逐渐显示出升高的ROS信号,大约20 min后达到峰值水平(图 3B-C,视频1)。 同时,在肿瘤细胞注射10 min后,部分tfRFP-C3-B16细胞中的C3 Ratio值逐渐降低,维持时间超过30 min,表明Caspase-3发生活化(图 3B-C,视频1)。
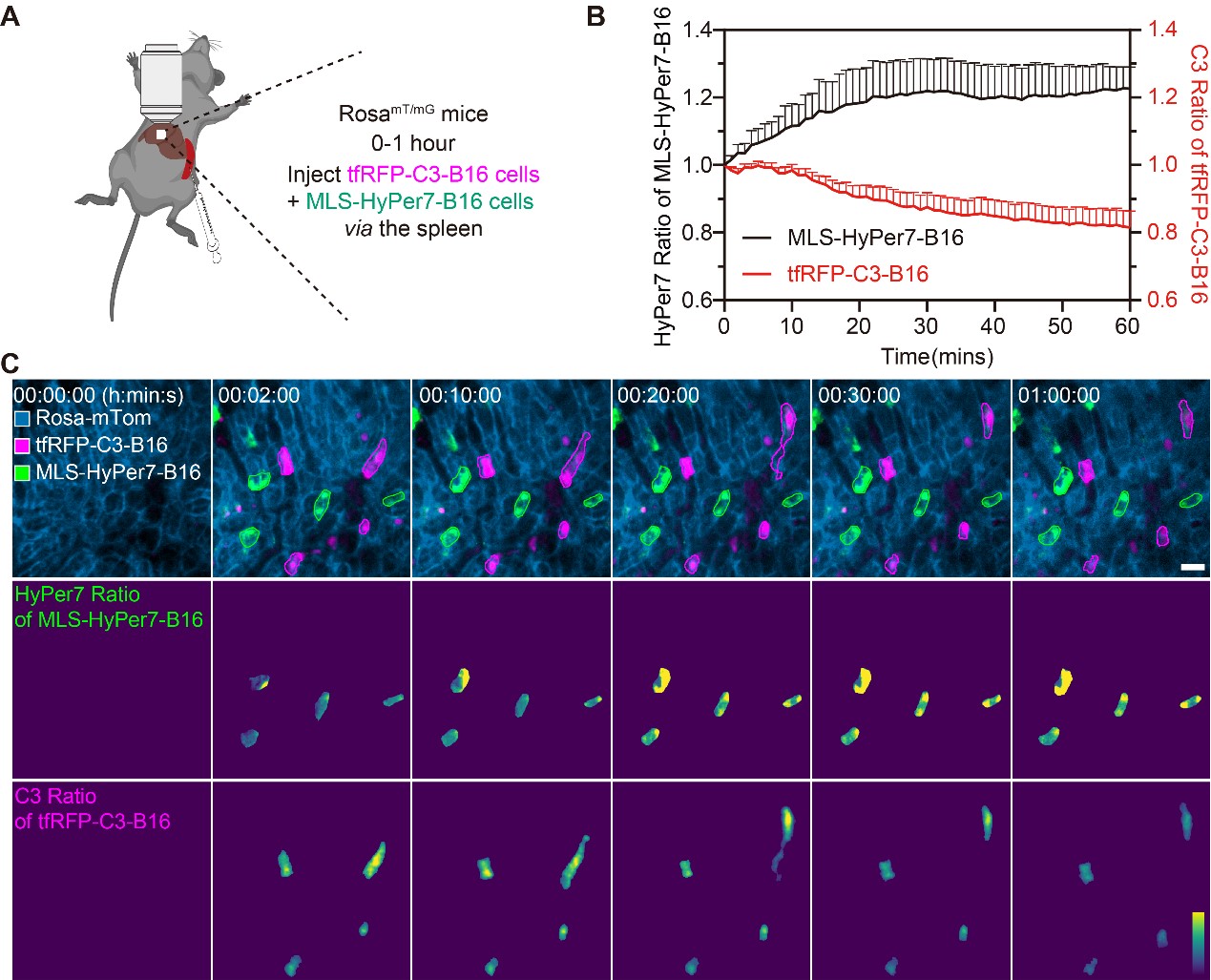
图 3. 肝脏活体成像观察转移到肝脏的肿瘤细胞ROS和Caspase-3分子信号变化。(A)肝脏活体分子成像示意图。(B)代表性的小鼠肝脏活体时间序列图像,上排为荧光图像,中间为分割的MLS-HyPer7-B16细胞的HyPer7 Ratio图像,下排为分割的tfRFP-C3-B16细胞的C3 Ratio图像。标尺:20 μm。(C)黑色曲线为B图中 MLS-HyPer7-B16 细胞中的HyPer7 Ratio变化曲线。
视频1:活体光学成像监测肿瘤转移 的早期分子事件。左侧图:MLS-HyPer7-B16细胞和 tfRFP-C3-B16 细胞混合注射后的活体成像图,第一行是荧光图,蓝色为Rosa-mTom标记的肝血窦,绿色为MLS-HyPer7-B16细胞,洋红为tfRFP-C3-B16细胞。第二行是MLS-HyPer7-B16细胞的HyPer7 ratio图,表示ROS信号强弱。第三行是tfRFP-C3-B16细胞的C3 ratio图,表示Caspase-3的活化水平。右侧图:肿瘤细胞tfRFP-B16细胞进入肝血窦早期的命运。免疫组小鼠体内可见tfRFP+微颗粒(0.75 ± 0.01 μm)形成,在未免疫小鼠体内可见tfRFP+囊泡(4.18 ± 0.54 μm)生成或细胞崩解荧光信号消失。
接下来研究者探究了升高的ROS与细胞膜损伤的关系。研究者使用500 μM浓度的柳氮磺胺吡啶(Sulfasalazine,SSA,作为ROS诱导剂)处理tfRFP-MLS-HyPer7-B16细胞(图 4A),验证了SSA能够诱导细胞ROS的升高(图 4B)。为了分析升高的ROS与细胞膜表面孔洞的关系,研究者使用原子力显微镜(AFM)观察了细胞膜表面形貌。结果显示,相比未处理的细胞,经过500 μM SSA处理后,tfRFP-B16细胞膜上有更多的孔洞(图 4C-D)。ROS诱导剂(SSA)与ROS抑制剂(N-乙酰半胱氨酸(NAC)或Ferrostatin-1)处理tfRFP-B16细胞24 h后,采用ELISA检测细胞培养基上清液中的tfRFP浓度。结果显示,与未处理组(3.25 ng/mL)相比,SSA组的tfRFP浓度(15.13 ng/mL)增加了4.66倍。SSA + NAC组(2.78 ng/mL)和SSA + Ferrostatin-1组(3.78 ng/mL)的tfRFP释放量相比未处理组没有显著增加(图 4E)。简而言之,这些结果表明,诱导肿瘤细胞的ROS升高能够导致细胞膜的通透性增加并引起胞内抗原的释放。使用AFM和共聚焦显微镜对tfRFP免疫小鼠血清处理的tfRFP-B16细胞同一区域进行成像(图 4F)。结果显示在tfRFP-B16细胞的细胞膜孔洞处存在tfRFP和IgG双阳性的抗原抗体复合物(图 4G)。这一结果表明细胞打孔引发释放的胞内抗原能够与抗体在细胞膜孔洞位点处结合形成抗原抗体复合物。
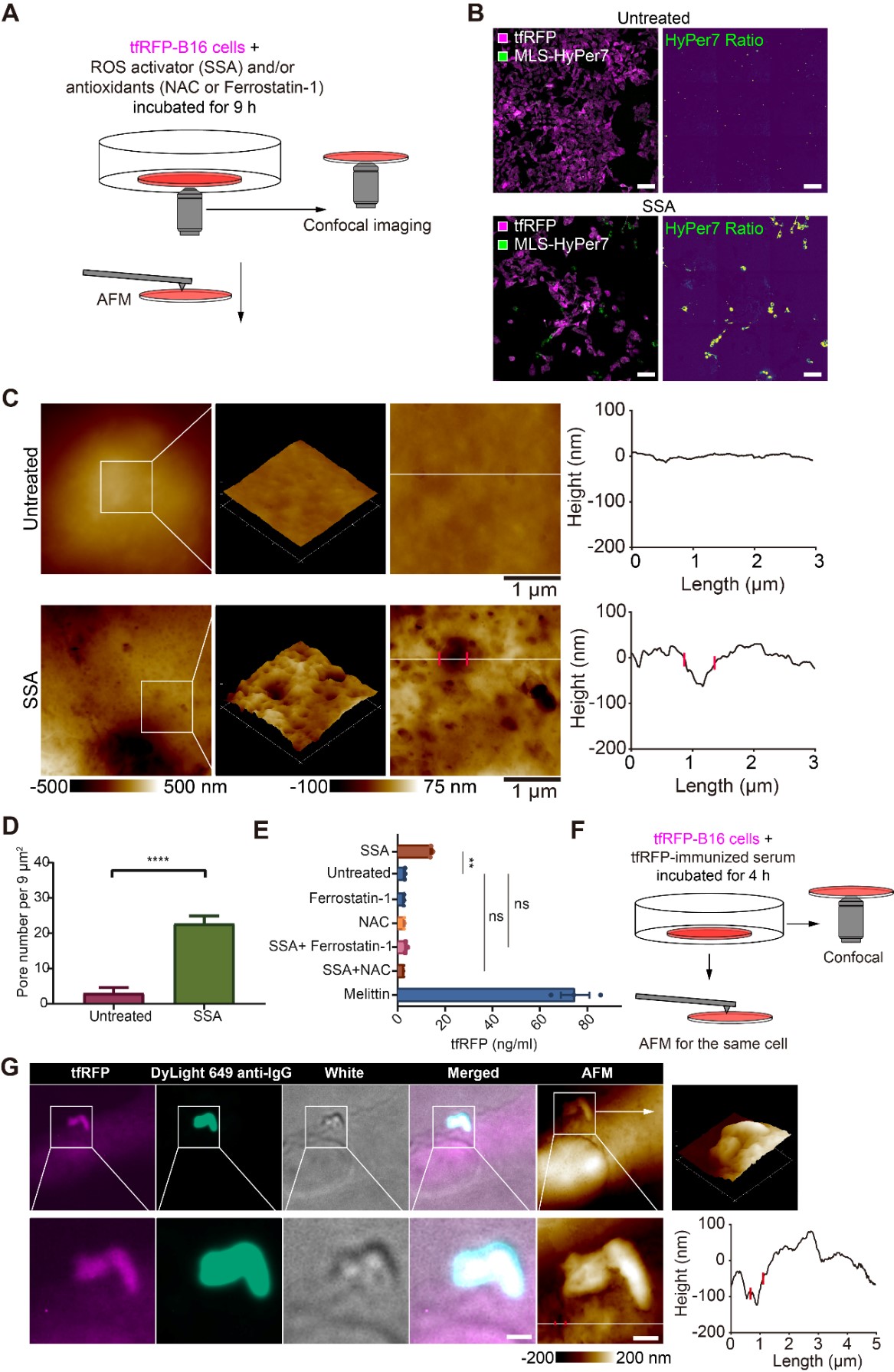
图 4. ROS是引发细胞膜打孔和胞内抗原tfRFP释放的关键因素。(A) tfRFP-B16细胞的共聚焦成像和原子力显微镜(AFM)成像示意图。(B)SSA处理9 h后的tfRFP-MLS-HyPer7-B16细胞共聚焦图,右图表征HyPer7 Ratio信号。图中各通道对比度保持一致,标尺:100 μm。(C)贴壁培养的tfRFP-B16细胞与500 μM SSA孵育9 h,经过多聚甲醛固定后,细胞膜形貌的原子力显微镜扫描图。图中白线穿过孔洞用于表征形貌,红点标记了孔的边缘,右图描绘了与白线对应的高度曲线。(D)对C图两组原子力扫描图中出现的孔洞密度进行的统计分析。数值展示为平均值 ± 样本标准误。(E) ELISA检测经不同药物处理后,tfRFP-B16细胞培养上清中tfRFP的浓度。数值展示为平均值 ± 样本标准误。ns:没有显著性差异,** P < 0.01(F)对同一tfRFP-B16细胞区域进行共聚焦成像和原子力显微镜扫描的实验示意图。(G)与tfRFP免疫小鼠血清孵育的tfRFP-B16细胞表面的共聚焦成像和AFM扫描成像。标尺:1 μm。
该研究后续通过原子力显微镜和ELISA等实验对GSDME-N和补体攻膜复合物在细胞膜打孔和胞内抗原tfRFP释放进行了检测,进一步验证了胞内抗原免疫清除肝转移瘤的机制。具体如下图5所示:本研究提出并验证了肿瘤细胞到达肝脏后通过活化ROS-caspase-3-GSDME通路引发肿瘤细胞膜打孔,胞内抗原通过肿瘤细胞膜表面孔洞释放,与胞外抗体结合形成抗原抗体复合物,抗原抗体复合物通过激活补体形成膜攻击复合物,膜攻击复合物在肿瘤细胞表面打孔,促进胞内抗原的进一步释放,形成更多的抗原抗体复合物。在这一过程中,中性粒细胞被招募到肿瘤区域对肿瘤细胞进行杀伤,死亡的细胞和抗原抗体复合物最终被肝脏巨噬细胞吞噬清除。总之,肿瘤细胞的ROS-caspase-3-GSDME通路活化是触发胞内抗原诱导的免疫清除肝转移瘤的关键步骤。
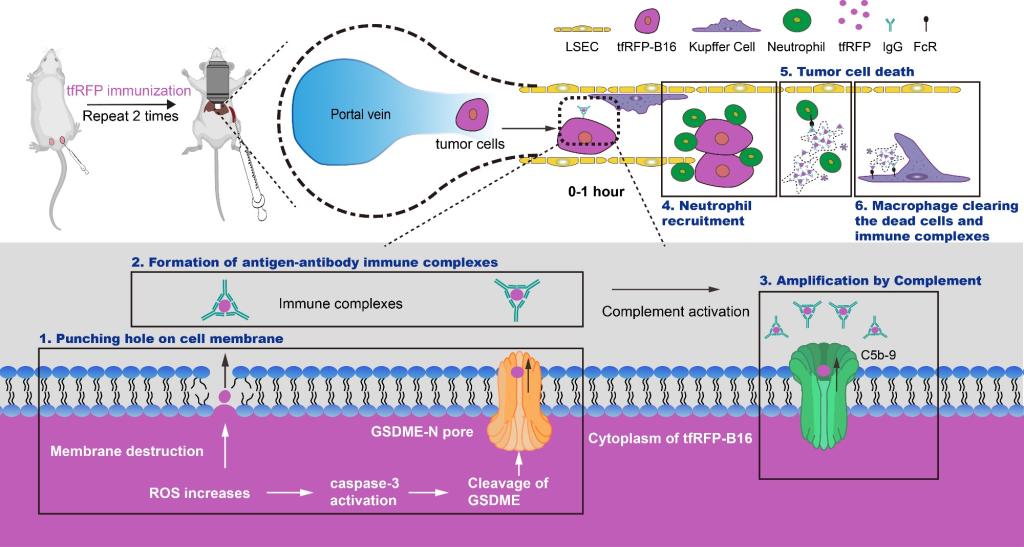
图 5. 胞内抗原诱导的免疫清除肝转移瘤的效应与机制示意图。
该研究得到国家重点研发计划(2017YFA0700403)、国家自然科学基金(91842305, 91842307, 81901691)、武汉光电国家研究中心创新基金和海南大学科研基金 (KYQD(ZR)20078)的资助。华中科技大学武汉光电国家研究中心张智红教授和祁淑红副研究员为文章的共同通讯作者,课题组研究生代博雷、章仁为文章的共同第一作者,海南大学骆清铭院士、课题组成员博士后刘征、研究生刘磊、张献、邓得强、张杰、本科生刘凡瑄和海南大学研究生许逸伦共同参与了相关工作。
原文链接:https://www.thno.org/v12p7603.htm